2025年10月7日,Nature Plants在线发表了云南大学、云南农业大学、中国科学院西双版纳热带植物园、加拿大多伦多大学、美国史密斯研究所国家自然历史博物馆、普洱学院多家单位共同完成的题为“Ginger genome reveals the SMPED1 gene causing sex-phase synchrony and outcrossing in a flowering plant”的研究论文(https://www.nature.com/articles/s41477-025-02125-3或https://rdcu.be/eJNGq)。该研究在已故著名植物进化生态学家李庆军教授二十余年系统研究“花柱卷曲性”(flexistyly)的基础上,以姜科(Zingiberaceae)山姜属(Alpinia)植物为研究材料,经历近十年的攻关,阐明了雌雄异熟的分子机制,鉴定出关键新基因AmSMPED1,并揭示其顺式调控与演化背景,为理解被子植物雌雄异熟的起源与维持机制提供了新的见解。

在被子植物中,雌雄异熟(dichogamy)是指同一朵花的雌蕊与雄蕊在不同时间成熟,这一机制能有效避免自交、促进异交,从而维持种群的遗传多样性。尽管自达尔文时代起,该现象就备受关注,但其分子调控机制长期未明,关键障碍在于多数物种的雌雄异熟类型是固定的,要么雌先熟(PG),要么雄先熟(PA),缺乏可用于遗传学解析的分离材料。山姜属独特的“花柱卷曲性”为破解这一难题提供了理想模型。雌先熟与雄先熟两种花型在自然种群中以稳定的1:1比例共存,且两种花型的雌雄器官每天都进行着精准同步且方向相反的节律性运动,有效地促进异花授粉。由此引出本研究的核心问题:究竟是哪一个遗传“开关”控制了这种异型雌雄异熟?其分子作用机理为何?
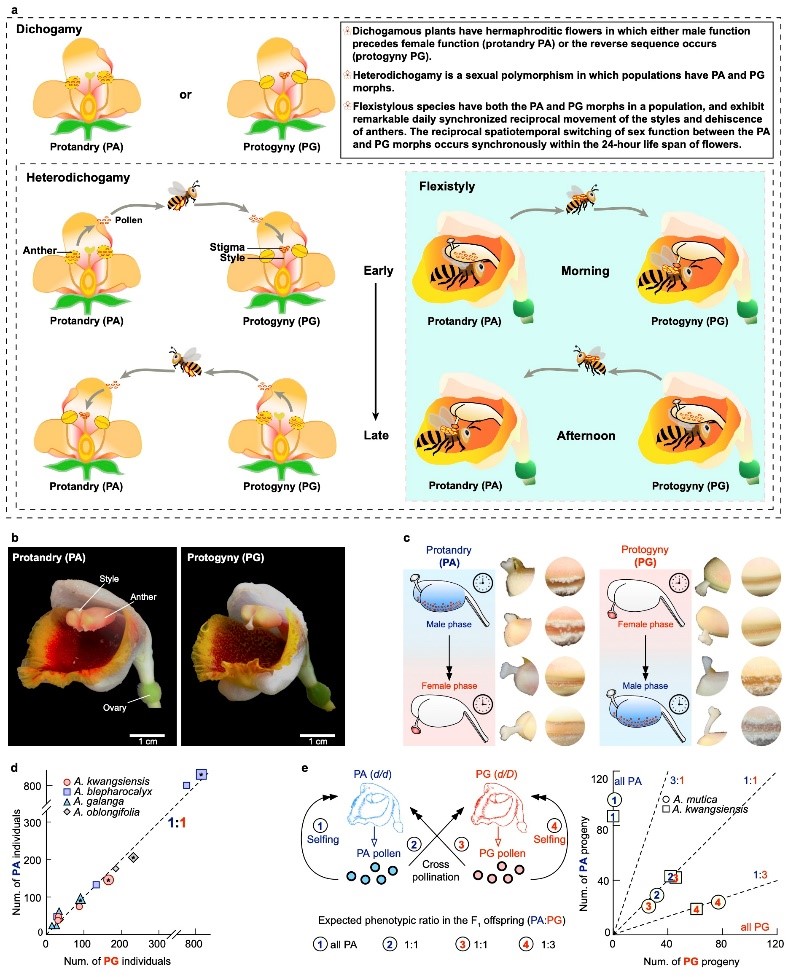
图1. 山姜属中雌雄异熟性状呈现孟德尔式遗传。a.雌雄异熟、异型雌雄异熟和花柱卷曲性关系;b. 马来良姜雌先熟和雄先熟的两种花型;c. 马来良姜两种花型开花当天互补的发育进程; d. 两种花型野外保持1:1比例共存;e. 两型杂交F1代符合孟德尔单基因显性模型。
研究首先通过对多个山姜属物种的自然种群调查与人工授粉实验,证实“花柱卷曲性”遵循经典孟德尔遗传规律。该性状由单一基因位点控制,决定雌先熟的等位基因为显性(D),决定雄先熟的等位基因为隐性(d),因此PG的基因型为杂合d/D,而PA为纯合d/d。在此基础上,团队构建了马来良姜(A. mutica)高质量单倍型基因组,并结合全基因组关联分析(GWAS),将控制该性状的遗传信号定位在8号染色体末端的“雌雄异熟决定区”(Dichogamy-Determining Region, DDR)。进一步分析显示,两种花型在该区域存在显著的基因组结构差异,DDR呈半合子(hemizygous)结构。与雄先熟相比,雌先熟在DDR区域多出一段包含完整LTR转座子的大片段插入,而雄先熟在相应位置表现为缺失。
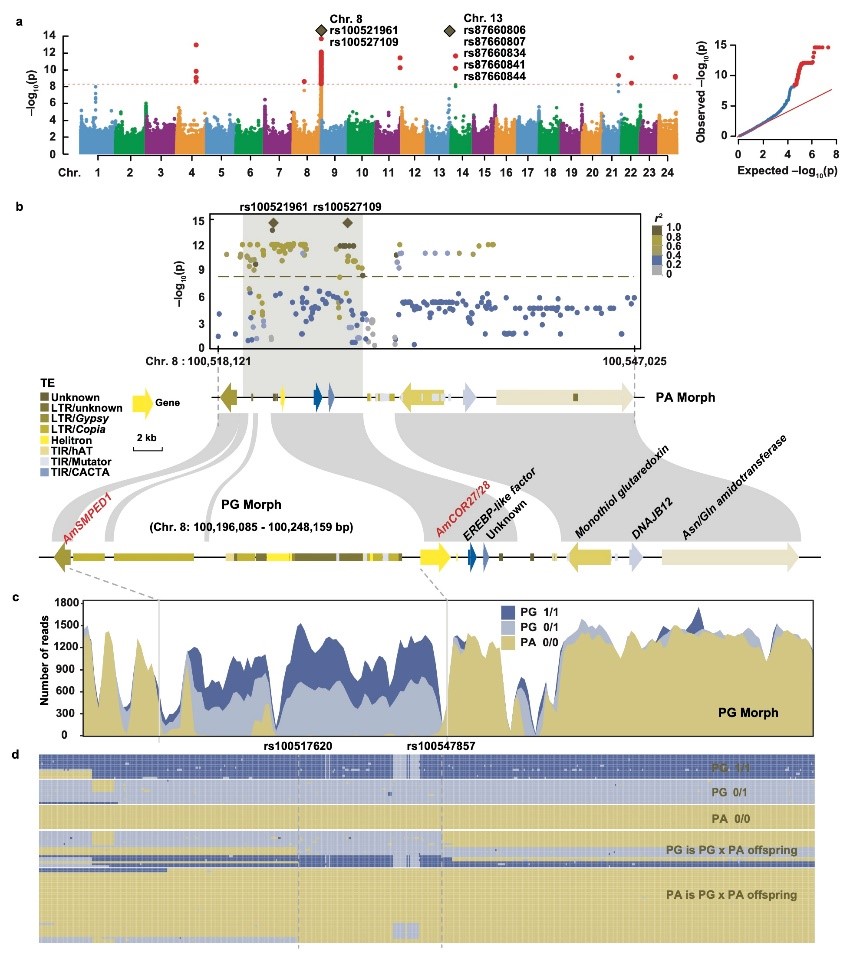
图2. 马来良姜基因组中鉴定“雌雄异熟决定区域”(DDR)。 a. 通过GWAS定位到雌雄异熟关键区域; b. 8号染色体末端连锁不平衡分析; c. 基于不同基因型的测序覆盖度曲线; d. 对实验群体的标记进行人工筛查。
DDR区域内含有多个候选基因。通过分析马来良姜雌雄器官在一天中不同时间点的基因表达节律,研究团队发现,只有一个此前未报道的新基因,其表达节律与花柱的每日运动和花药的开裂时间高度同步。团队将其命名为SMPED1(STYLE MOVEMENT and POLLEN EARLY DISPERSAL 1)。为最终确认AmSMPED1(Am为马来良姜中拉丁名简写)是否具有调控花柱运动和花粉囊开裂的功能,研究团队先通过反义寡核苷酸技术瞬时下调AmSMPED1在花柱中的表达,结果花柱的节律性运动随之停止。其次,团队利用模式植物拟南芥验证了AmSMPED1与花药开裂的关系。将马来良姜的AmSMPED1基因导入拟南芥,可使其花药比野生型提前数小时开裂;这一现象在拟南芥中AmSMPED1基因的两个同源基因也被观察到,分别过表达AtSMPED1/2过表达株系提前于野生型散粉,同时降低AtSMPED1/2的表达则会延迟开裂。这些证据共同证实,AmSMPED1是同时调控雌蕊运动和雄蕊成熟时间的多效性基因。
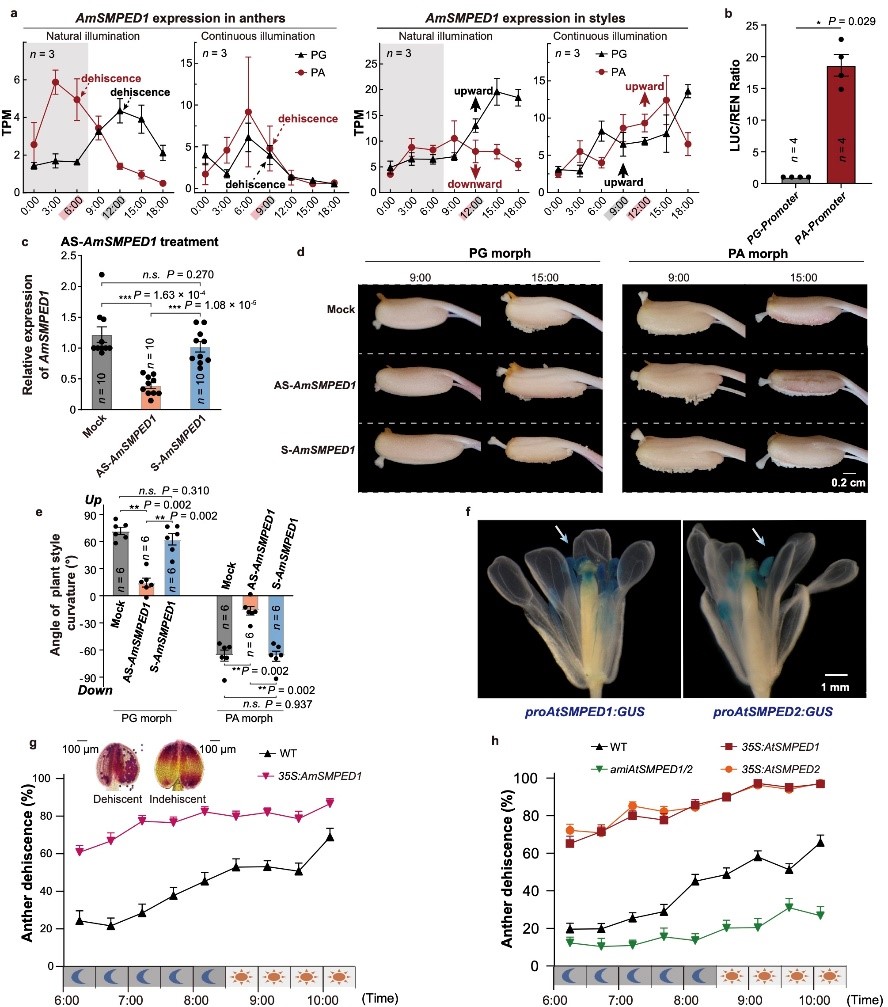
图3. 验证SMPED1是一个多效性基因。 a. 在自然光照与连续光照下AmSMPED1的基因表达与花柱运动和花粉囊开裂的时序相对应; b. 两种花型启动子活性存在差异; cde. 反义寡核苷酸下调AmSMPED1的表达并抑制花柱运动; fgh. AmSMPED1基因在拟南芥中有两个同源基因特异性的在花粉囊中表达,并且它们都正向调控花粉囊开裂。
对等位差异与调控机制的进一步剖析显示,两种花型中AmSMPED1的蛋白编码序列仅存在一个同义突变。等位基因特异表达与启动子活性实验表明,显性等位基因D的启动子活性低于隐性等位基因d,在雌先熟d/D的背景下由d等位基因主导表达。上述结果将两种花型差异明确指向顺式调控改变,LTR转座子的插入重塑了AmSMPED1的表达调控程序,根本性地改变了其表达节律,从而产生并维持了两型花在时间上的精确分工。
在演化层面,姜目中存在一个保守的祖先DDR;在姜科约∼32.5 Mya的全基因组复制后,DDR拷贝分化为DDR I与DDR II。DDR II是表型分化的关键,祖先DDR II上发生的LTR插入(在马来良姜中可追溯至∼4.06 Mya)产生了PG花型。
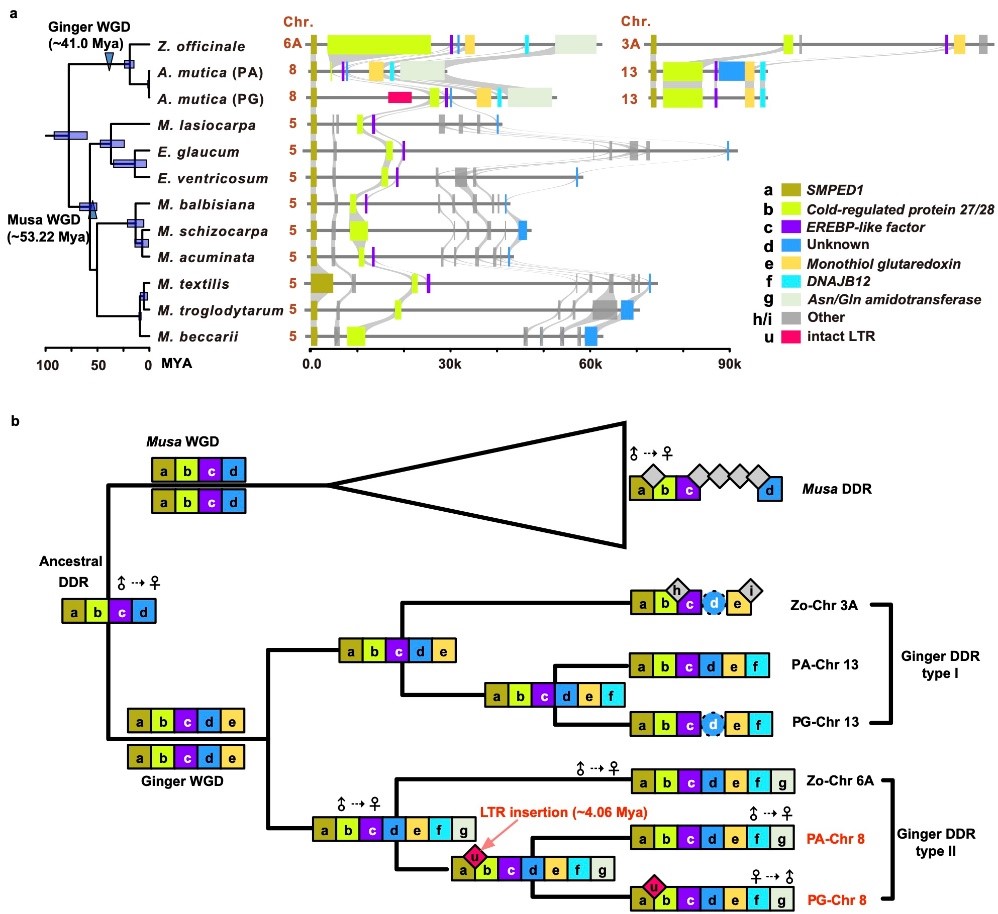
图4. DDR的起源。 a. 构建芭蕉科与姜科的系统发育树,并对各物种的DDR进行比对,展示分化时间、全基因组加倍事件及共线同源关系; b. 依据基因获得与丢失事件重建祖先DDR,揭示其组成及演化轨迹。
不同于DDR,SMPED1基因几乎在所有被子植物中均可找到,其蛋白结构与功能预测高度保守,提示这是一个参与被子植物生殖过程的保守基因。据此提出,SMPED1原本与花药开裂相关,通过LTR介导的半合子结构与顺式调控改变,在山姜属中演化出雌先熟与雄先熟两套调控程序,从而形成并维持“花柱卷曲性”。
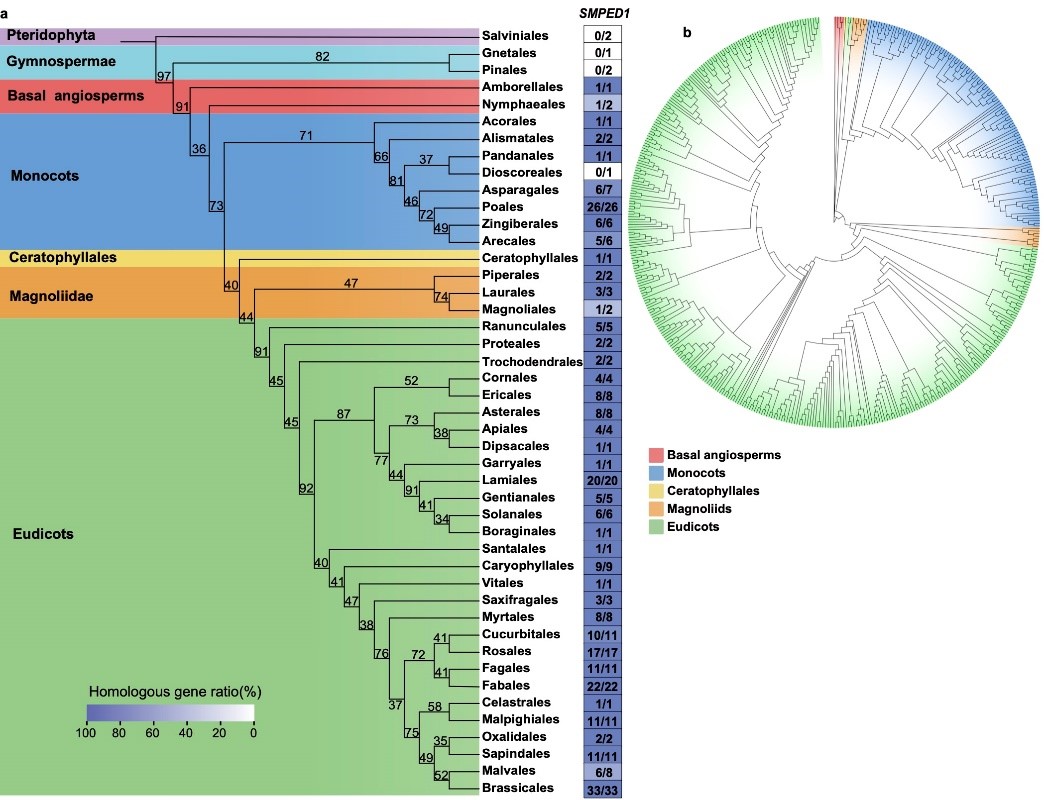
图5. SMPED1的植物谱系演化。 a. SMPED1在被子植物中的分布; b. 解析SMPED1基因树。
综合而言,这项研究确定花柱卷曲性由单一孟德尔位点控制,首次清晰地描绘了一个复杂的雌雄异熟系统的分子机制;发现并功能验证了AmSMPED1是驱动雌性(花柱运动)和雄性(花药开裂)功能的多效性基因;阐明了DDR在姜目中的起源与结构演化;并提供了SMPED1在被子植物中的广泛保守性证据。本研究不仅鉴定了一个控制植物雌雄功能同步的新基因,也阐明了雌雄异熟现象的起源与维持,指出由转座子介导的基因组结构变异和调控重塑在其中扮演了关键角色。这一发现对于理解植物性系统的起源和演化具有深远意义,并可能为未来作物育种提供新的理论依据和技术靶点。
云南大学赵建立研究员、黄奥丹博士后、彭晓昶博士后,云南农业大学董扬教授、段胜常博士为论文共同第一作者。云南大学赵建立研究员、韩佳嘉副研究员,云南农业大学陈玮教授,加拿大多伦多大学著名生物学家Spencer C.H. Barrett教授为论文共同通讯作者,云南大学李庆军教授为论文最后一位作者。该工作得到了云南省科学技术厅与云南大学“双一流”建设联合项目、国家重点研发计划课题、国家自然科学基金、云南省“青年人才计划”、云南大学研究生科研创新基金资助。
李庆军教授在Nature期刊报道“花柱卷曲性”(flexistyly)后(Li et al. Nature 410: 22),迅即引发学界广泛关注。此后,他与团队成员从生态学、生态生理学、进化生物学等方面持续深入探索花柱卷曲性的机制。然而,在非模式植物中揭示其分子机制,难度远超预期。约十年前,李庆军教授牵头联合相关领域专家协同攻关,研究逐步取得突破。遗憾的是,就在项目如火如荼推进之际,这一现象的发现者、本文工作的引领者李庆军教授,于2022年12月1日不幸辞世。科研之路漫长,成果来之不易,却未能与开创者共同见证,令人深痛。本文的发表,寄托对李庆军教授的深切缅怀,也铭记他在揭示植物生殖策略演化方面的卓著贡献。