近日,云南大学生态与环境学院生物多样性研究院的植物演化生态学团队在一区TOP期刊Communications Biology在线发表题为Standing genetic variation and introgression shape the cryptic radiation of Aquilegia in the mountains of Southwest China的研究论文,揭示了西南山地耧斗菜属物种隐存辐射谱系间形态的相似性与快速分化的演化机制,对解析中国西南山地及周边地区的生物多样性具有重要意义。
隐存辐射是多个形态上相似但遗传上存在显著差异谱系的集合,也是生物多样性中尚未被充分认识的重要组成部分。团队以毛茛科(Ranunculaceae)耧斗菜属(Aquilegia L.)在我国西南山地的无距耧斗菜复合群为研究系统,通过野外调查采集了该区域内23个自然群体,并利用全基因组重测序技术,对西南山地耧斗菜属物种的系统发育关系、群体遗传结构、渐渗和分子进化等方面进行了分析,解析了隐存辐射的演化过程和机制。
基于系统发育关系和群体结构的结果,团队将西南山地耧斗菜的三个形态学物种界定为11个进化谱系,每个形态学物种内都包含3至4个隐存谱系(图1)。该界定结果也得到了地理、气候、繁殖生态学以及花色证据的支持。群体历史相关分析结果表明系统发育不一致主要受到渐渗和谱系分选不完全(incomplete lineage sorting)的影响。对于形态学物种间的固定分化位点的筛选和分析则进一步显示在甘肃耧斗菜和直距耧斗菜的谱系中观察到的形态关联位点的起源时间都早于谱系形成时间,支持谱系分选不完全对于相似形态关联位点的塑造(图2)。全基因组窗口的遗传多样性和结构的关联分析结果支持该复合群的基因组岛的快速形成主要受到固有遗传变异分选模型的驱动(图3)。此外,对谱系特异的分化位点的遗传来源和驱动力的解析结果,表明大部分隐存谱系的适应性分化位点都受到了非姐妹谱系间渐渗的影响(图3b和3c)。
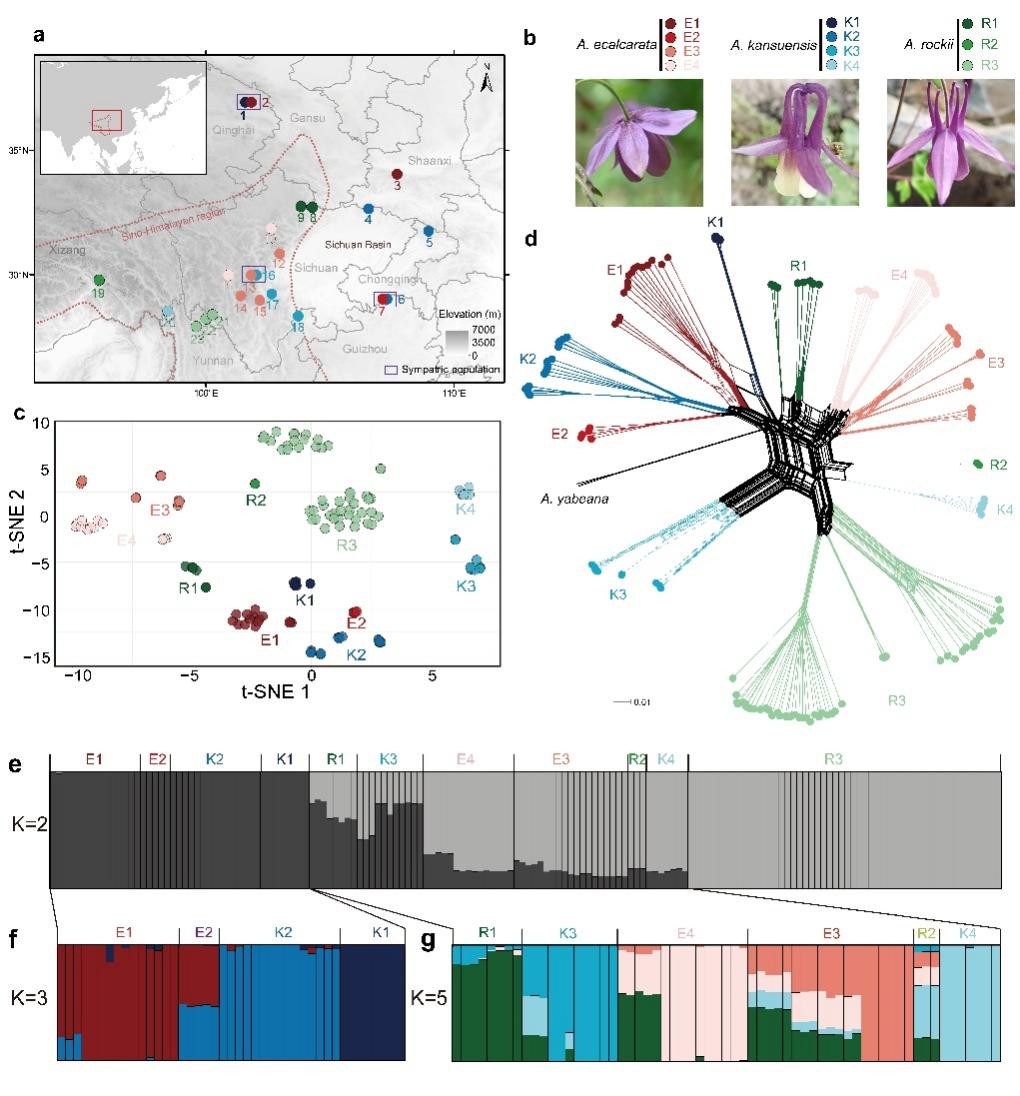
图1 西南山地耧斗菜属物种的遗传结构和进化关系。(a)本研究中23个群体采样分布图。(b)形态物种的花部照片(c)基于全基因组不连锁SNP数据集的t-sne 结果。(d)基于距离的系统发育网络结果。(e-g)基于STRUCTURE软件推断的遗传结构。
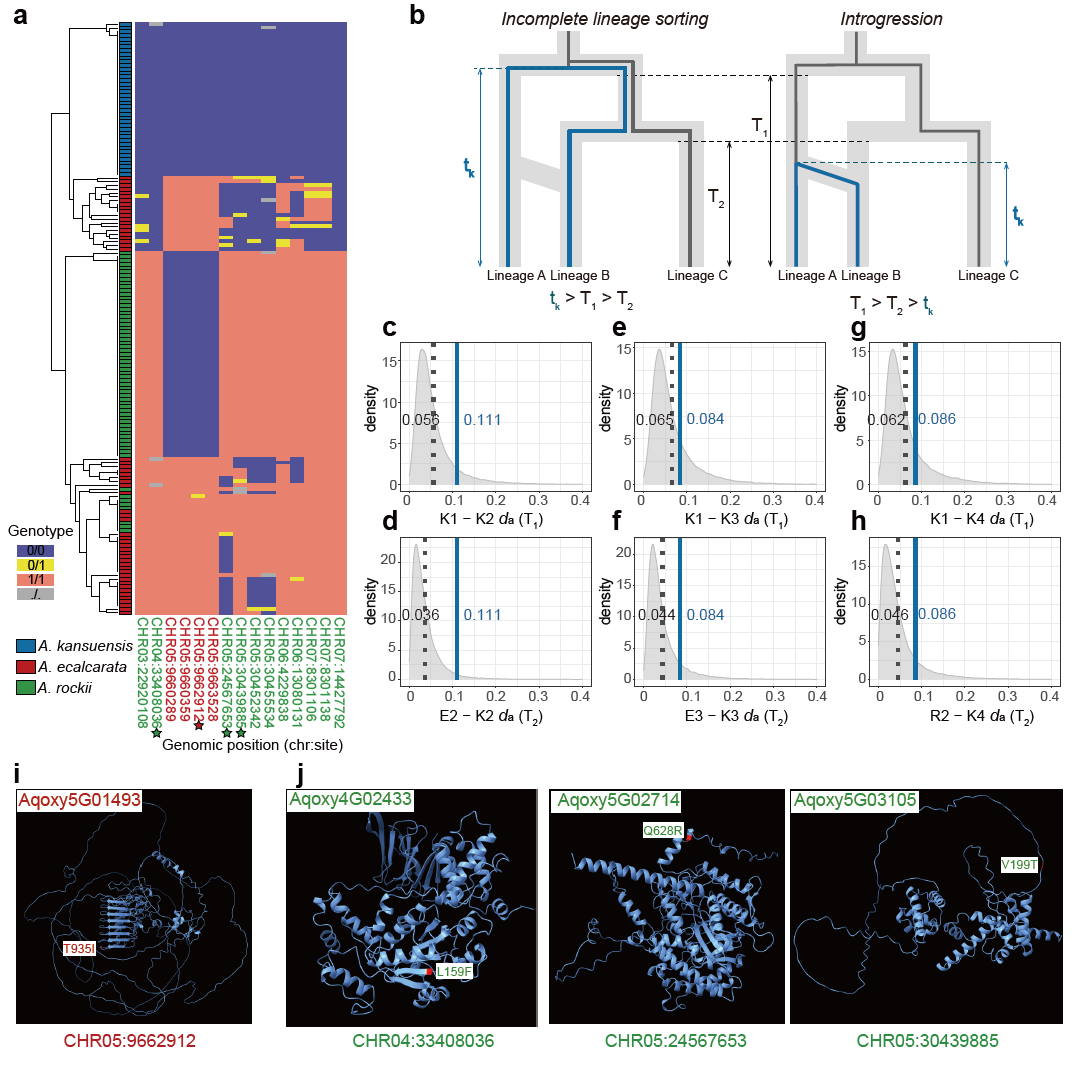
图2 甘肃耧斗菜固定分化位点的演化分析结果。(a)固定分化SNP位点的单倍型聚类。星号标记为位于基因编码区域的位点,均为非同义突变,同时固定位点在基因组上呈现聚集,符合单次起源的旁系演化(collateral evolution)特征。(b)固定分化位点演化假设的图示,进一步用于解释甘肃耧斗菜中固定位点单次起源的两种演化假设,即不完全谱系分选(左)和渐渗(右)。(c-h)甘肃耧斗菜隐存谱系和姐妹谱系之间的形态关联位点和谱系分化程度的比较。蓝色直线代表固定分化位点的分化程度,而黑色虚线代表所有位点的分化程度,均以平均da 来代表。(i和j)Alphafold预测的四个关联基因的蛋白质结构。
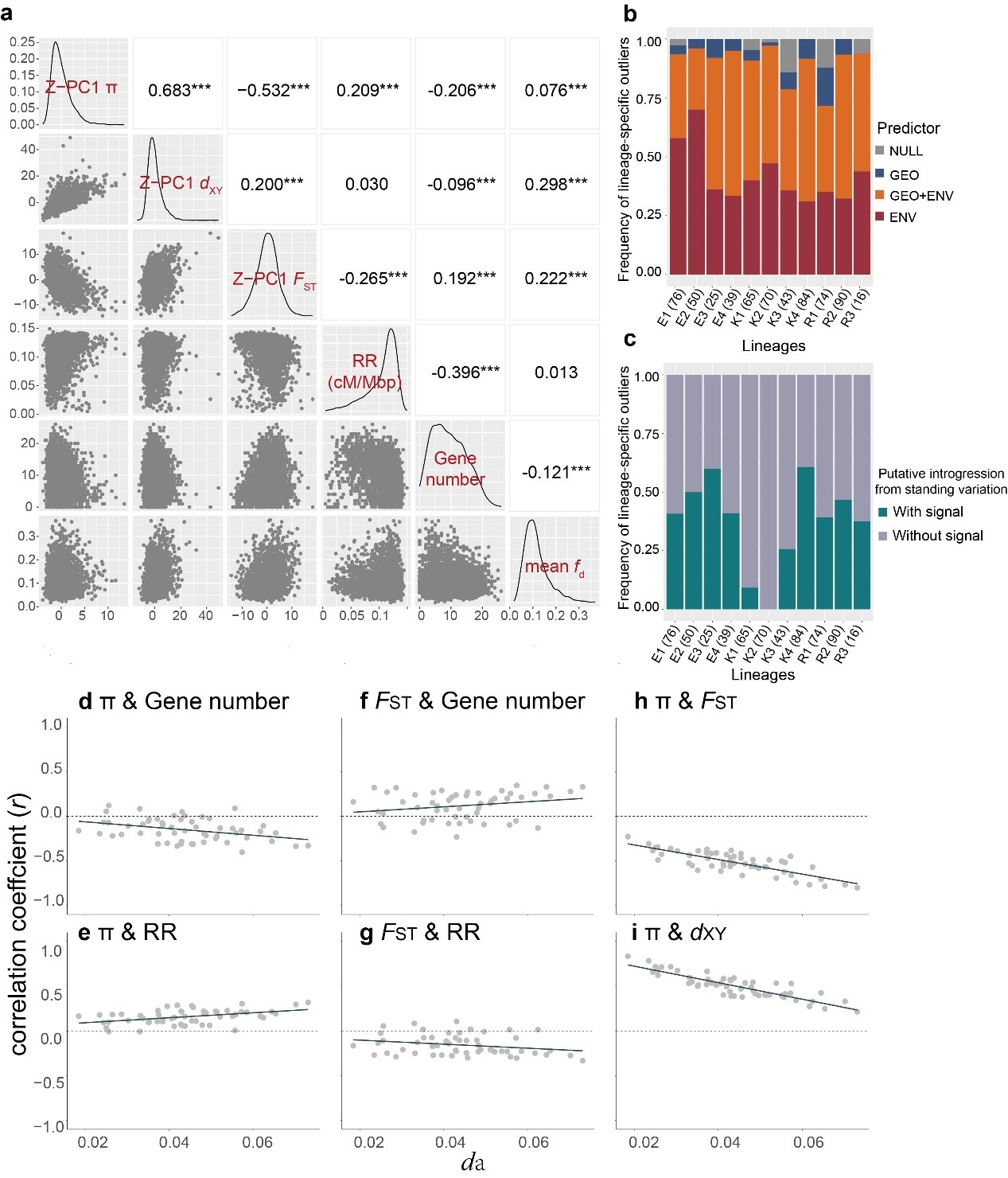
图3 基因组结构与遗传多样性之间的相关性。(a)基于100 kb非重叠窗口内估计的PC1 π、PC1 FST、PC1 dxy、平均fd、基因密度和重组率的成对相关矩阵。PC1,第一主成分。***为p < 0.001;**为p < 0.01;*为p < 0.05;无星号为p > 0.05。(b)基于DIC权重的谱系特异分化位点与环境和地理关联分化模式的比例。(c)非姐妹谱系间固有遗传变异渗入受体谱系异常位点的比例。(c)和(d)中括号内的数字表示每个谱系中检测到的谱系特异分化位点(Z-PBS > 2和Z-ΔFST >2)的数量。(d-i)55次谱系之间遗传变量比较的相关性(以灰点代表)与分化时间(以da平均值代表)的趋势变化。
该研究在厘清西南山地耧斗菜物种的系统发育关系和群体结构的基础上,进一步明确了谱系间快速遗传分化和相似形态的演化机制。其中,不完全谱系分选是塑造甘肃耧斗菜和直距耧斗菜相似形态演化的重要原因,而隐存谱系间的快速遗传分化则受到了固有遗传变异和渐渗的共同塑造。该研究解析了固有遗传变异和渐渗在隐存辐射演化中的重要作用,也为理解植物隐存辐射的演化机制提供了经典案例。
云南大学博士后彭俊楚为第一作者,张志强副研究员为通讯作者,中山大学何子文教授参与了本工作。该研究得到了国家自然科学基金(32271693, 32300203)和云南省基础研究计划(202405AC350111, 202501AS070067)的支持。
原文链接:https://www.nature.com/articles/s42003-025-08120-w